
不是人眼没有演化出看见紫外线的能力,而是人类的祖先“退化”掉了看见紫外线的能力。
其实对于有胎盘类、有袋类哺乳动物以及它们组成组成的兽类哺乳动物而言,能看到紫外线才是祖先特征,后期看不到紫外线是各个支系特化形成的衍征,同样包括人类在内的灵长类动物看不到紫外线,是“抛弃”了祖先的技能。
在具体分析之前,我们先解释一下动物是怎样看到紫外线的。
动物是如何看到紫外线的?
我们都知道人类的眼睛可以感受三种颜色,红色、绿色和蓝色,即光学三原色,这只是最简单的说法。
实际上,我们是通过视锥细胞中的视紫红质(视蛋白 + 视黄醛)来感受颜色的,人类具有三种视锥细胞,每种视锥细胞中都具有单一的视紫红质,所以人类的眼睛中具有三种感受色觉的视紫红质,它们对应的电磁波(光)最大吸收波长(λmax)分别为 564–580nm,534–545nm 和 420–440nm(不同人类个体不同,所以为范围值),对应的颜色大概就是黄绿色、绿色和蓝紫色,波长上则被称为长波(L)、中波(M)和短波(S)。
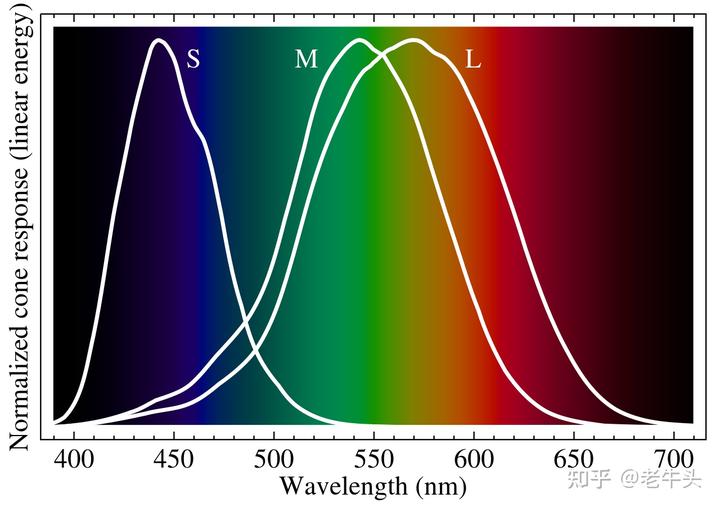
其中,能够感受短波长的视紫红质的视蛋白被称为短波敏感视蛋白(short wavelength-sensitive opsin1,OPN1SW/SWS1)。在人类中,OPN1SW 的λmax 为 420-440nm,而我们对于紫外线的定义为 400-10nm,人类的 OPN1SW 对于波长小于 400nm 的电磁波的敏感度几乎为 0,所以人眼看不到紫外线。
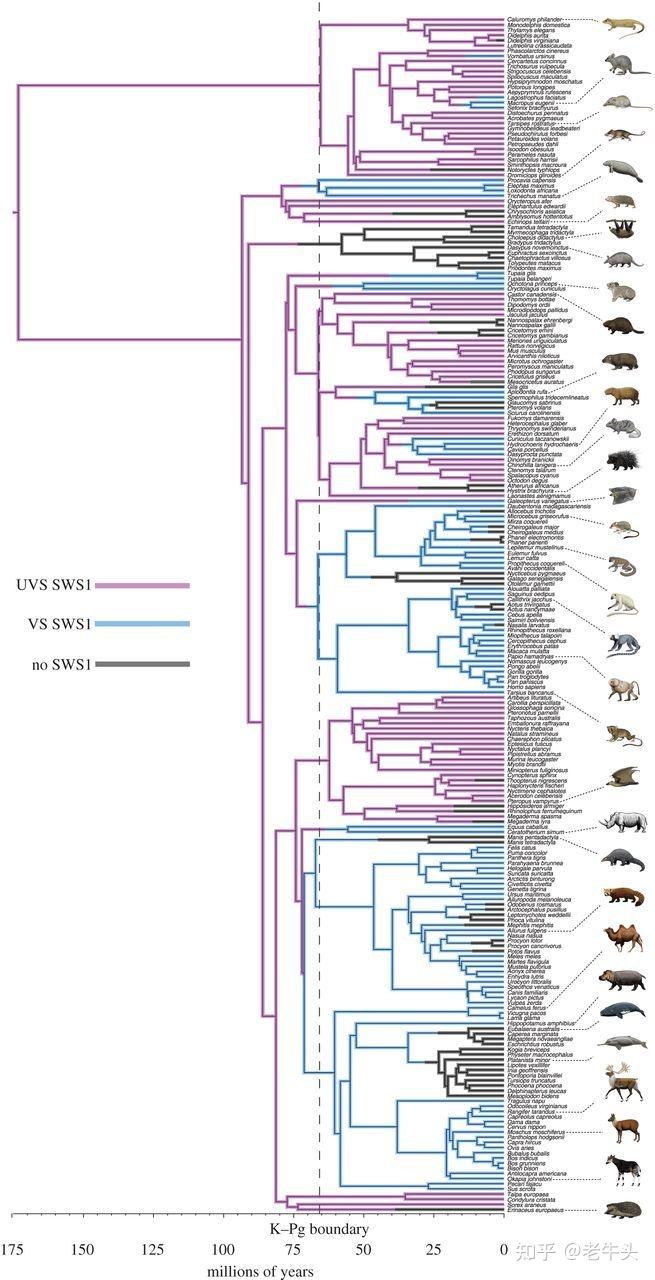
而除了人以外的其他哺乳动物,如果它们的 OPN1SW 的λmax 小于 400nm 的话,它们的短波视锥细胞就可以感受到紫外线,紫外线对于它们来说就是可见光。这些哺乳动物主要包括有袋类(袋鼠、树袋熊、袋獾等)、啮齿目(各种鼠)、翼手目(蝙蝠)和真盲缺目(鼹鼠、鼩鼱和刺猬)。
不过,除了眼睛可以直接看见紫外线的哺乳动物外,一些哺乳动物比如麋鹿的 OPN1SW 可以感受到紫外线但并不直接形成清晰的像[1],对于它们来说,发出或者反射紫外线的物体上可能覆盖着一层深蓝色的阴影,而不是具体的颜色。[2]
为什么不同哺乳动物的 OPN1SW 感受到的光线波长不一样呢?
对于哺乳动物来说,根据 OPN1SW 的λmax 所在范围,可以分为紫外光敏感(UVS)和紫光敏感(VS),其中 UVS 型的的 OPN1SW 的λmax 的范围为 350-400nm,而 VS 型的 OPN1SW 的λmax 的范围为 400-450nm,很显然人类的 OPN1SW 属于 VS 型。而哺乳动物的 OPN1SW 的λmax 与其氨基酸序列有关,OPN1SW 上的一些氨基酸位点的变化会影响其λmax.
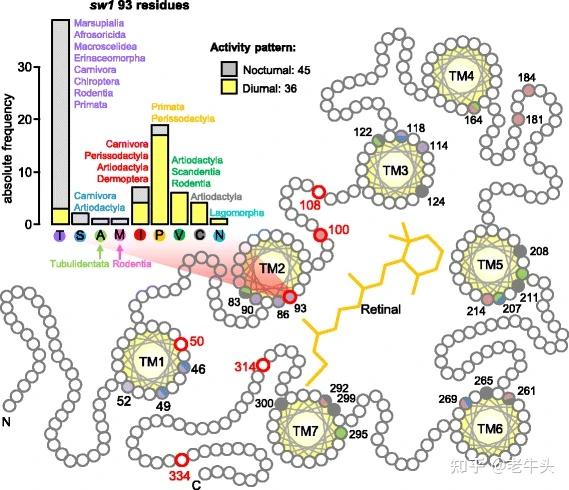
比如 OPN1SW 的第 86 号氨基酸,在有胎盘类哺乳动物中其祖先状态为苯丙氨酸,如果这个氨基酸替换为其他氨基酸(比如酪氨酸、丝氨酸或缬氨酸),那么哺乳动物的 OPN1SW 就极有可能从 UVS 型变为 VS 型。
但对于灵长类来说,93 号位点的氨基酸更为重要。在灵长类中,OPN1SW 的 93 号氨基酸从苏氨酸向脯氨酸的转变很可能是其 VS 型 OPN1SW 起源的关键因素。
在对马达加斯加指猴(aye-aye,Daubentonia madagascariensis)的研究中,研究者发现,其 OPN1SW 的 86 号氨基酸为苯丙氨酸(祖先型),但它的 OPN1SW 是 VS 型而不是 UVS 型,但如果把它的 93 号氨基酸转为苏氨酸,则 OPN1SW 的λmax 从 406nm 减小至 371nm,从 VS 型转为 UVS 型。[3]
包括人类在内的所有现生灵长类,其 OPN1SW 的 93 号位点的氨基酸均为脯氨酸,这是灵长类 OPN1SW 为 VS 型的一个基础。
同时通过对现生所有灵长类最近共同祖先 OPN1SW 的氨基酸序列进行祖先状态重建,并与灵长类的近亲啮齿类进行对比,研究者发现灵长类与啮齿类的 OPN1SW 氨基酸序列在涉及到影响 OPN1SW 的λmax 的氨基酸点位上只有两个点位有差异,93 号和 114 号,93 号上文中已经说到了,在灵长类中从苏氨酸转变为脯氨酸,而 114 号通过研究,则是从啮齿类所代表的祖先状态 - 丙氨酸,转变为了甘氨酸,这说明除了 93 号氨基酸,114 号氨基酸对于灵长类 VS 型 OPN1SW 的起源也有重要的影响,两者的变化共同导致了人类的灵长类祖先看不到紫外线。[4]
在有胎盘类、有袋类和兽类(有胎盘类和有袋类组成的类群)哺乳动物中,UVS 型的 OPN1SW 是祖先状态,所有 VS 型的 OPN1SW 都是通过基因突变导致的氨基酸改变产生的。[5]
那么,为什么灵长类要“退化”掉看见紫外线的能力呢?
在中生代,现生哺乳动物的祖先主要在夜间和黄昏活动,这种习性可能起源于炎热的三叠纪,可能要早于哺乳动物的起源。在三叠纪,哺乳动物的祖先体型偏小,内温性,维持体温的恒定,白天炎热的气温对于没有很好散热机制的它们来说可能是致命的,甚至会杀伤它们的雄性生殖细胞[6],所以哺乳动物的祖先选择了在更凉爽的夜间和黄昏进行活动,这种习性一直存在到白垩纪末大灭绝。
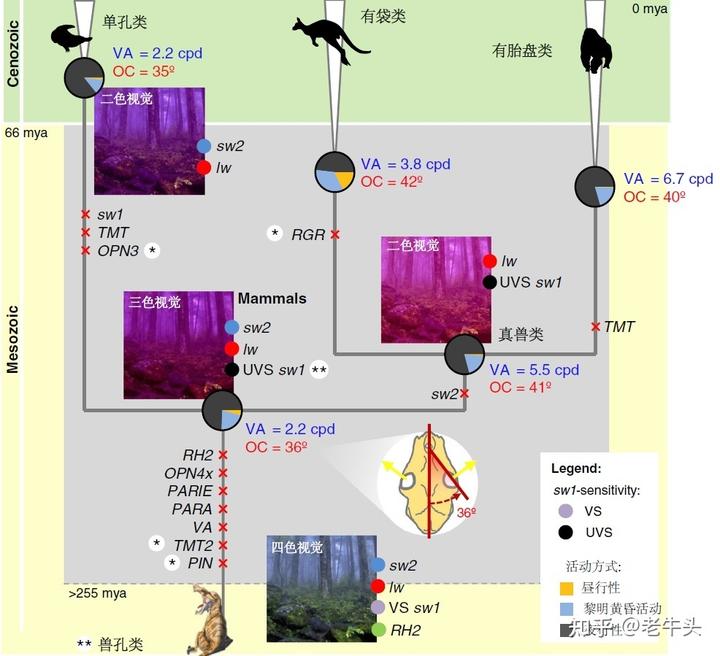
在夜晚和黄昏活动,理论上应该不太需要看见紫外线,因为这时候的紫外线要比白天更弱,那为什么哺乳动物在中生代的祖先拥有紫外线视觉呢?
很遗憾,这个问题至今没有答案。我个人的理解是这是对于祖先特征的一种存留,也许哺乳动物的犬齿兽类甚至兽孔类祖先本身就具有紫外线视觉,这对于转化为夜行性的哺乳动物来说没有改变(发生演化)的需求,所以就保留了下来。由于哺乳动物与现生其他羊膜动物(爬行类和鸟类)的亲缘关系很远,我们无法知道现生哺乳动物在二叠纪和三叠纪祖先的 OPN1SW 演化的过程。
但关于为何有胎盘类哺乳动物的 OPN1SW 从 UVS 型向 VS 型转变,研究者有较为成熟的认识。
在白垩纪末大灭绝后,随着陆地生态系统中大量捕食者的灭绝,哺乳动物可以在白天活动,这时强烈的紫外线可能会造成视网膜的损伤,于是日间活动的哺乳动物的晶状体对于紫外线的透过率会下降,减弱进入眼球的紫外线以保护视网膜。[4]
同时,随着白垩纪末大灭绝后,众多有胎盘类哺乳动物类群的体型普遍增大,它们眼球的长度也随之增加,这也减少了紫外线的透过率。
为了更好的感光和成像,OPN1SW 的λmax 需要从紫外光区移到紫光区,这便是对白天活动的、体型增大的哺乳动物的选择压力,于是使得 OPN1SW 的λmax 变大的基因突变可以更好地保留下来,最终形成了 VS 型的 OPN1SW.
所以,在现生有胎盘类哺乳动物中,除了啮齿目、翼手目和真盲缺目等主要在夜晚和黄昏活动的小型哺乳动物以外,大部分大型的日行性哺乳动物都的 OPN1SW 都是 VS 型,这也是灵长类失去看到紫外线能力的原因。
总结
总之,人类看不到紫外线,不是因为人眼没有进化出看到紫外线的能力,而是因为包括人类在内的所有现生灵长类的祖先“退化”掉了看到紫外线的能力,这是灵长类从祖先的夜行性习性转向日行性习性以及体型变大所必须做出的适应。而这种适应是通过基因突变改变短波敏感视蛋白(OPN1SW)的氨基酸位点上的氨基酸,从而改变其最大吸收波长(λmax)做到的。



















